|
…ий вопрос, который требовал выяснения: какова обучаемость аппортировке у собак, обладающих стремлением носить предметы до всякого обучения. Три собаки с подобным поведением (Чанг, Альфа, Джульбарс) были дрессированы аппортировке. Эти собаки чрезвычайно легко дрессировались этому приему. Две из них, Джульбарс и Чанг, с первого дня занятий начали подносить предметы. Третья собака, Альфа, хватала предметы с первого дня обучения, однако подносить их по команде начала со второго занятия. Дальнейшим этапам обучения дрессировались только две собаки (Альфа, Джульбарс). Обе эти собаки очень легко выдрессировались всем четырем этапам. Альфе понадобилось всего 10 занятий (наименьшее число занятий из всех дрессированных собак). Джульбарс выдрессировался на 15-й день занятий. Таким образом, собаки, имеющие стремление носить предметы, обучаются и самой аппортировке и всему, что с ней связано, чрезвычайно легко. По данным Шама, среди охотничьих собак имеется специально выведенная в Англии порода — ретриверы — для аппортировки убитой дичи. Собаки этой породы — великолепные аппортировщики, обладающие, помимо этого свойства, еще умением совершенно не мять подносимую дичь, качеством, особенно ценимым охотниками у ретриверов. Ретривер был выведен путем скрещивания некоторых пород охотничьих собак и Лабрадора, породы собак, которая еще в начале XIX столетия была ввезена в Англию и известна охотникам как очень хороший аппортировщик, который, согласно Хагену, должен был передать эти способности ретриверу. Данные о большой легкости, с которой ретривер дрессируется аппортировке, мы находим у Дарвина. Эти данные полностью согласуются с нашим выводом о наличии у собак наследственно обусловленного различия в быстроте выработки такого условного рефлекса, как подношение хозяину тех или других предметов. Может возникнуть вопрос: не является ли данное поведение собак, обучающихся легко аппортировке и имеющих стремление носить предметы, результатом общих свойств их нервной системы, связанных с повышенной способностью к выработке условных рефлексов вообще, и не является ли быстрая обучаемость аппортировке лишь одним из проявлений этого свойства? Полученные мною данные говорят против такого предположения. Одна из упомянутых выше собак, Дже-Амур, обладавшая стремлением носить предметы, была исследована мной в условно-рефлекторной камере по обычной слюнно-секреторной методике. Работа в камере показала, что Дже-Амур вырабатывал условные рефлексы со средней скоростью. Так, например, первый условный рефлекс на метроном, 120 ударов в минуту (отставление 30 с), начал вырабатываться на 6-м опыте (25–30 сочетаний) и достиг значительной прочности на 10-м опыте (58–64 сочетаний)… Второй условный рефлекс на звонок (совпадающий) выработался после 20 сочетаний. Дифференцировка оказалась хорошей, давая частые 0. По типу нервной деятельности собака может быть отнесена к сангвиникам с довольно хорошим тонусом коры головного мозга. Никаких особенных отклонений в условно-рефлекторной деятельности не было обнаружено. Таблица 2. Быстрота выработки условного рефлекса «садиться» у собаки, обладающей склонностью к аппортировке, по сравнению с собакой, не обладающей этим свойством
Примечание: В обоих случаях рефлекс выработался на 9-м опыте.
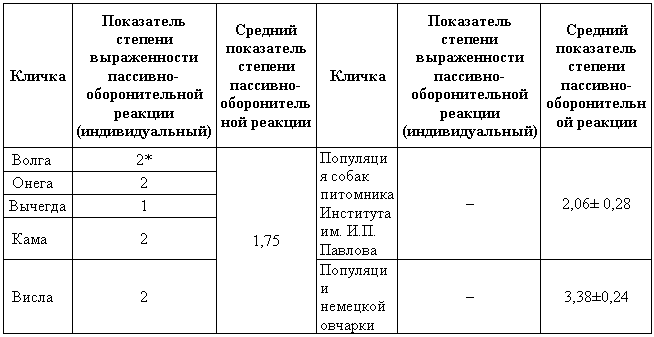
Вторая собака, Чанг, обладавшая стремлением носить предметы и начавшая почти без всякого обучения аппортировать, была дрессирована приему садится по команде «сидеть» с точной реакцией хода обучения. В табл. 2 приведены данные хода обучения этой собаки по сравнению с собакой, не обладающей стремлением носить предметы. Безусловный раздражитель: нажим рукой на крестец и рывок за поводок. Как видно из табл. 2, выработка двигательных условных рефлексов у обеих собак происходила довольно сходно. Несколько более быстрый ход выработки рефлекса у Чанга не настолько велик, чтобы его можно было принять во внимание. Таким образом, у собак, обладающих стремлением к аппортировке, вырабатываются условные рефлексы, не связанные с ней, в пределах обычной нормы. Условные рефлексы, связанные с аппортировкой, вырабатываются у них исключительно легко. Итак, приведенные нами данные показывают следующее. У собак имеется значительная изменчивость в быстроте и легкости выработки двигательного условного рефлекса аппортировки предметов. В крайней форме выражение этого свойства проявляется в стремлении собак без предварительного специального обучения носить в зубах различные предметы. Данное свойство обусловливается генотипически. Стремление к аппортировке и легкость выработки условных рефлексов, связанных с ней, по-видимому, не являются выражением повышенной легкости и быстроты выработки у таких собак всех рефлексов. Данное свойство является специфической особенностью собаки, проявляющейся на фоне нервной деятельности, в остальном обычной. Формирование оборонительного поведения у собакВ формировании оборонительного поведения тесно переплетаются влияния внешних и внутренних факторов, что и создает возможность использовать это формирование для освещения вопроса о зависимости между врожденными и приобретенными формами поведения. Как показали опыты Майорова (1933), собаки, выращенные в изолированных условиях содержания, отличаются значительно выраженной пассивно-оборонительной реакцией (трусостью). Согласно Павлову, у таких собак сохраняется щенячий рефлекс «естественной осторожности», который у собак, живущих в условиях свободы, затормаживается еще в щенячьем возрасте при знакомстве со всем многообразием внешнего мира. Отсюда, казалось бы, следовало, что пассивно-оборонительная реакция является чисто приобретенным свойством, возникающим в ответ на определенные условия воспитания. Однако в дальнейшем мной было показано, что в возникновении данной формы поведения большую роль играет и наследственность. Оказалось, во-первых, что при скрещивании двух собак, отличающихся преобладанием пассивно-оборонительной реакции, подавляющее число потомков отличается такой же формой поведения; во-вторых, что при скрещивании таких собак с собаками, не отличающимися преобладанием данной формы поведения, процент потомков с пассивно-оборонительной реакцией гораздо меньше, чем в первом случае; в-третьих, что при скрещивании собак, не отличающихся пассивно-оборонительной реакцией, большинство потомков также не отличается данной формой поведения (Крушинский, 1938). На первый взгляд казалось, что имеются две сходные по своему проявлению оборонительные реакции: врожденная и приобретенная. Однако дальнейшие данные показали, что изолированные условия воспитания вызывают преобладание пассивно-оборонительного поведения в тесной зависимости от генотипа собаки. Оказалось, что различные породы собак обладают весьма различной степенью выражения пассивно-оборонительной реакции при их воспитании в условиях изоляции. На рис. 2 сопоставлены данные о степени выраженности пассивно-оборонительной реакции у эрдельтерьеров и немецких овчарок, воспитывающихся в условиях изоляции (питомник). Как видно из рис. 2, у эрдельтерьеров процент собак с пассивно-оборонительной реакцией невелик; у немецких же овчарок пассивно-оборонительной реакцией отличается подавляющее большинство особей.
Рис. 2. Пассивно-оборонительная реакция при воспитании в условиях изоляцииПо оси ординат — процент особей с пассивно-оборонительной реакцией, по оси абсцисс — степень выраженности пассивно-оборонительной реакции; 1 — эрдельтерьеры; 2 — немецкие овчарки.
Рис. 3. Пассивно-оборонительная реакция при свободном воспитанииОбозначения те же, что и на рис. 2.
Таблица 3. Пассивно-оборонительная реакция доберман-пинчеров (все самки, полная изоляция) по сравнению с пассивно-оборонительной реакцией собак других пород, воспитанных при частичной изоляции
* Показатели 1–4 обозначают возрастающую степень выраженности пассивно-оборонительной реакции. Методика определения описана подробно в моих статьях: «Роль наследственности и условий воспитания в проявлении и выражении пассивно-оборонительной реакции у собак» и «Наследование пассивно-оборонительного поведения (трусости) в связи с типами нервной системы у собак».
О том же говорит и следующий опыт: пять щенков доберман-пинчера (породы собак, не склонных к проявлению данной формы поведения) были воспитаны нами в Институте эволюционной физиологии и патологии высшей нервной деятельности им. И. П. Павлова в условиях полной изоляции (забитый доверху досками вольер). Пассивно-оборонительная реакция проявилась у этих собак в весьма незначительной степени.
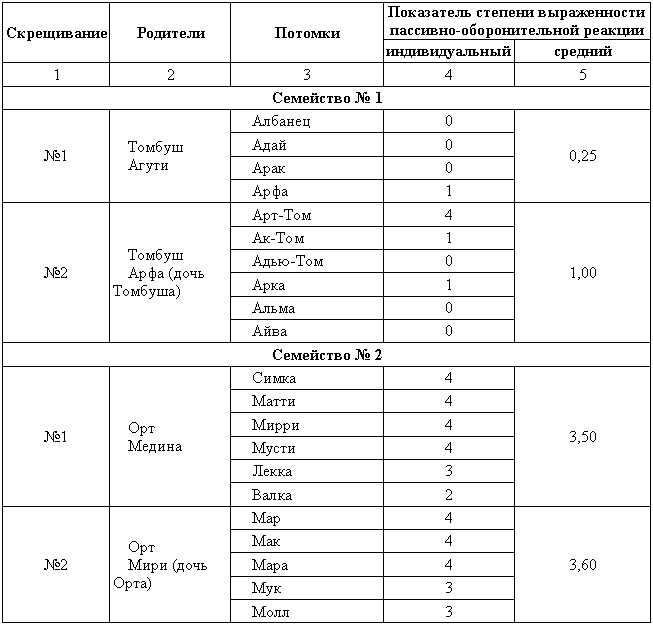
Как видно из табл.3, несмотря на то, что доберман-пинчеры воспитывались в условиях, весьма благоприятных для проявления пассивно-оборонительной реакции (полная изоляция), это свойство оказалось у них менее резко выраженным, чем у собак других пород, воспитывавшихся в условиях меньшей изоляции. Значение генотипа для проявления пассивно-оборонительной реакции у собак ясно видно и из коэффициента корреляции между степенью выраженности данной реакции у братьев и сестер, воспитанных в одних и тех же условиях частичной изоляции питомника Института эволюционной физиологии и патологии высшей нервной деятельности им И. П. Павлова (r=+0,54±0,06). Далее, резко различной оказалась выраженность пассивно-оборонительной реакции у двух семейств собак, воспитанных в одних и тех же условиях этого питомника (табл. 4). Эти данные свидетельствуют о большой роли генотипа в проявлении и выражении пассивно-оборонительной реакции, возникающей у собак в ответ на изолированные условия воспитания. Данная реакция в своем проявлении и выражении находится в тесной зависимости как от условии воспитания, так и от генотипа собаки. Возникает вопрос: может ли пассивно-оборонительная реакция у собак, имеющих генотипическую наклонность к ее проявлению, проявиться не только в изолированных условиях, но и при воспитании в обычных условиях свободы. Можно привести данные, которые это подтверждают. Во-первых, немецкие овчарки — собаки, обладающие большей наклонностью, чем эрдельтерьеры, к проявлению пассивно-оборонительной реакции в условиях изоляции, и при обычном воспитании на свободе все же резче проявляют эту реакцию, чем эрдельтерьеры (рис. 3). Во-вторых, собаки, принадлежащие к различным семействам, различаются степенью выраженности у них пассивно-оборонительной реакции при воспитании в условиях как изоляции, так и свободы. Из табл. 5 видно, что существует явное соответствие между степенью выраженности пассивно-оборонительной реакции у собак сходных генотипов при их воспитании в условиях свободы и изоляции. Собаки, обладающие генотипом, способствующим значительному выражению пассивно-оборонительной реакции при изолированных условиях воспитания, отличаются и при свободном воспитании некоторой пассивно-оборонительной реакцией. Таким образом, имеется различная норма реагирования у различных собак к проявлению пассивно-оборонительной реакции. С одной стороны вариационного ряда находятся собаки, обладающие генотипом, который обусловливает малую наклонность к данной реакции поведения. Такие собаки проявляют пассивно-оборонительную реакцию только при строго изолированных условиях воспитания, да и то в незначительной степени. Таблица 4. Степень выраженности пассивно-оборонительной реакции у собак двух семейств, воспитанных в одних и тех же условиях частной изоляции (питомник Института эволюционной физиологиия патологии высшей нервной деятельности им. И. П. Павлова)
С другой стороны вариационного ряда находятся собаки, обладающие генотипом, способствующим исключительно легкому проявлению пассивно оборонительной реакции. Такие собаки проявляют эту форму поведения даже при свободном воспитании. Между этими крайними полюсами помещается непрерывный ряд особей, обладающих различными нормами реагирования в проявлении пассивно-оборонительной реакции. Таблица 5. Соотношение между степенью выраженности пассивно-оборонительной реакции потомков трех различных производителей при свободных и изолированных условиях воспитания
Приведенный анализ формирования двух различных реакций поведения показал сходные явления. Во-первых, наличие значительной изменчивости в индивидуально приобретенном поведении, возникающем в ответ на сходные внешние воздействия. Эта изменчивость определяется различным генотипом животных. Во-вторых, наличие непрерывного ряда переходов» от особей, у которых определенное поведение формируется под ведущим влиянием внешних факторов, до особей, у которых данное поведение возникает под ведущим влиянием внутренних, генотипических факторов. В-третьих, возможность исключительно легкого, почти без всякого предварительного опыта, формирования определенного поведения у тех особей, у которых имеются соответствующие генотипические предпосылки к его возникновению. В то же время у других особей такое поведение возникает только после длительного индивидуального опыта или очень сильного воздействия внешних факторов. Проведенное исследование выработки условного рефлекса аппортировки и формирования оборонительного поведения у собак принесло ряд фактов, подтверждающих выдвинутые нами представления о соотношении между индивидуально приобретенными и врожденными реакциями поведения. Приведенные факты указывают, что при возникновении в популяции индивидуально приобретенной биологически полезной формы поведения могут создаваться условия для действия естественного отбора в направлении замены роли «внешних» факторов, определяющих возникновение этого поведения, «внутренними» факторами. Процесс такой замены можно понимать как фиксирование в популяции врожденного, инстинктивного поведения, развивающегося под действием отбора из материала, выявляемого индивидуально приобретенным поведением животного. Рассмотрим на примерах исследованных нами реакций поведения ход этого, с нашей точки зрения неизбежного, процесса. Предположим, что для какой-нибудь популяции собак (или других животных) биологически выгодно выработать условный рефлекс ношения или подноски тех или других предметов и особи этой популяции выработают соответствующий условный рефлекс. Быстрота образования этого рефлекса будет весьма различной у отдельных особей. Наличие изменчивости в этом отношении ясно видно из приведенного выше материала. Те особи, у которых этот рефлекс вырабатывается быстрее, будут иметь преимущество в борьбе за существование. Это поведет к положительному отбору соответствующих особей; отобравшиеся особи передадут по наследству своим потомкам соответствующие генотипические предпосылки. Начнется отбор из поколения к поколению особей, обладающих наклонностью к выработке этого рефлекса. Кривая изменчивости в популяции будет от поколения к поколению сдвигаться в сторону ускорения образования этого условного рефлекса. Так как крайние представители вариационного ряда, наиболее легко вырабатывающие рассматриваемый условный рефлекс, отличаются, как мы видели выше, генотипически обусловленным поведением даже без предварительного обучения носить во рту те или другие предметы, то естественный отбор поведет к постепенному увеличению процента таких собак, пока все особи популяции не будут обладать этим свойством поведения. Как только это осуществится, мы сможем сказать, что произошла замена внешних факторов, игравших ведущую роль в формировании данного поведения, внутренними факторами, т. е. условно-рефлекторное поведение у особей этого вида заменилось безусловно-рефлекторным инстинктивным поведением. Совершенно такой же процесс будет происходить с пассивно-оборонительной реакцией. Предположим, что какая-нибудь популяция собак одичала. Воспитываясь в условиях, где щенки не видят людей, вырастающие собаки сохранят рефлекс естественной осторожности, который проявится в виде пассивно-оборонительной реакции. Степень ее выраженности, обусловленная различным генотипом отдельных особей, будет весьма различной. Предположим далее, что люди начнут уничтожать этих одичалых собак. В первую очередь уничтожат тех особей, которые обладают наименее резко выраженной пассивно-оборонительной реакцией. Начнется положительный отбор собак, проявляющих пассивно-оборонительную реакцию в наиболее резкой форме в ответ на данные условия существования, и потомки этих собак даже при домашнем воспитании сохранят наследственно «зафиксированное» поведение. Основной направляющей причиной развития врожденного безусловно-рефлекторного поведения из индивидуально приобретенного является, таким образом, естественный (или искусственный) отбор по наследственным наклонностям к возникновению тех или других индивидуально приобретаемых реакций поведения. Теоретические соображенияАнализ приведенных выше фактов показывает, что при формировании различных реакций поведения переход ведущей роли от внешних факторов к внутренним может обусловливаться биологической необходимостью. Биологическая необходимость замены внешних факторов, определяющих поведение, внутренними, врожденными факторами, проливает, как нам кажется, свет на вопрос о путях образования целесообразности инстинктивного поведения животных. Поражающая нас целесообразность, приспособленность врожденных инстинктивных форм поведения к тем условиям, в которых живет животное, делается понятной, если рассматривать данную форму поведения как эволюционирующую по пути, проложенному наследственно зафиксированным индивидуальным опытом. Все многообразные пробы и ошибки, посредством которых каждое животное наилучшим образом приспосабливается к условиям жизни, создавая материал для действия отбора, могут оказать тем самым формирующее влияние и на поведение потомков. Одной из причин целесообразности инстинктивности поведения животных может явиться биологическая необходимость замены (под влиянием отбора) внешних факторов формирования поведения внутренними. Хотя между условным и сложно-безусловным рефлексом и имеется глубокая качественная разница, однако эволюция сложно-безусловного рефлекса из «наследственно-фиксирующегося» условного рефлекса могла идти по пути чисто количественного превращения. Путь этой эволюции нам представляется в следующем виде. Животные, попадающие в изменившиеся условия существования и выработавшие определенные условные рефлексы — привычки, приспосабливающие их к новым условиям жизни, могут передавать своим потомкам некоторые наклонности к выработке этих условных рефлексов. На первом этапе это может происходить путем теснейшего переплетения передающихся путем подражания традиций и результатов начавшегося отбора. Ллойд Морган приводит следующий факт: «после того как через шотландские болота проведены были телеграфные проволоки, тетерева, ударявшиеся о проволоки, на первых порах умирали в больших количествах. Но спустя год или два это вымирание прекратилось». По этому поводу он задает вопрос: «Унаследовали ли молодые птицы привычку летать, избегая проволок? Или эта привычка была сообщена им с помощью традиций?» Мы полагаем, что на этот вопрос надо ответить так: возможно, что и традиция, и начавшийся отбор особей, научившихся наиболее легко избегать провода и воспринимать самую эту традицию, шли рука об руку, тесно переплетаясь друг с другом. На этой стадии наследственная готовность к приобретению определенных навыков может быть еще и столь незначительной, что если исчезают новые условия, то сейчас же исчезает и начавшее приспосабливаться к ним поведение животных. Так, Вагнер указывает, что «фазаны, которых держали в полуневоле, отводя им определенные лесные участки и ставя кормушки, тетерева, которых из поколения в поколение держали в неволе, приобретали новые навыки вместо тех, которым следовали, руководясь инстинктом, но, вернувшись к свободной жизни, те и другие теряли приобретенное, и прежние инстинкты у них восстанавливались в своем первоначальном виде» (Вагнер, 1925. С.56). Более длительное пребывание животных в новых условиях может закреплять наклонности к выработке специфических, необходимых в данных условиях навыков. На этой стадии наследственного фиксирования приобретаемых индивидуальным опытом навыков возможно развитие специальных наклонностей к более легкой выработке определенных рефлексов, помогающих животным в их борьбе за существование. С подобным случаем легкой выработки определенного условного рефлекса мы встречаемся, очевидно, у гончих собак, обладающих наклонностью «сваливаться» в кучу по первому же сигналу, «ибо в течение сотен поколений особи, не усваивающие себе требуемых навыков, убивались доезжачими» (Вагнер, 1913). Дальнейшая стадия «фиксирования» индивидуально-приобретенных реакций поведения может выражаться уже не только в легкости выработки определенных условных рефлексов, но и в том, что некоторые черты данного индивидуального рефлекса будут проявляться в последующих поколениях без всякого предварительного индивидуального опыта. Индивидуальный опыт в этом случае будет лишь развивать и усиливать те черты поведения, которые уже оказываются наследственно закрепленными отбором. Сюда относится, как мы полагаем, появление натурального рефлекса когда собака исключительно быстро, «с места» начинает вырабатывать условный рефлекс на незнакомый ей до этого времени запах съедобного вещества (Цитович, 1911). К этой же группе наследственно фиксирующихся условных рефлексов должно быть отнесено, возможно, и поведение гончих собак. Характерное поведение гончей собаки выражается в том, что собака с лаем преследует по следу зверя. Для развития данного поведения необходима так называемая «нагонка», т. е. особая тренировка. Однако характерное поведение гончих собак может проявляться и в отсутствии внешнего раздражителя, обычно его вызывающего: зверя и его следа. Проявление подобного поведения мне удалось наблюдать на стае очень породистых молодых гончих собак, еще никогда не бывших на охоте. Будучи выпущенными в большой отгороженный лесной выгул, некоторые из этих собак проявили совершенно характерное поведение гончей собаки, гоняющей зверя по следу. Они нюхали землю и с совершенно типичной для гончих собак манерой с лаем «гнали» отсутствующего и никогда ими не виданного зверя. Притом эта мнимая охота носила весьма упорный характер. Собаки эти никогда до этого не были на охоте и никаких следов в выгуле не было. Щенки эти воспитывались с месячного возраста под моим наблюдением в вольерах около нашей лаборатории. Отсутствие следов в выгуле я специально проверял по только что выпавшему снегу. В то время этот факт произвел на меня большое впечатление. В самом деле, при отсутствии раздражителя (зверя или его следа) у собак, которые никогда до этого не были на охоте, но только принадлежали к известной линии охотничьих собак, проявилась типичная реакция в отсутствие специфического внешнего раздражителя. Следовательно, аппарат, необходимый для данного поведения, созрел и готовность к нему настолько оформилась, что даже в отсутствии внешнего раздражителя эта созревшая готовность к реакции начинала неудержимо проявляться, обнаруживая результат того длительного искусственного отбора, которому в сотнях поколений подвергались предки современных высокосортных гончих собак. Подобные проявления инстинктивных реакций при отсутствии специфических внешних раздражителей приведены и в литературе. Лоренц описывает, например, поведение скворца, выращенного в искусственных условиях, который в отсутствие мух производил мнимую на них «охоту» со всеми характерными для нее деталями поведения. Проявление подобного поведения при отсутствии специфического внешнего раздражителя Лоренц назвал «реакцией вхолостую». По его мнению, проявление подобных реакций характерно для «чистых инстинктов», порог раздражения которых делается настолько низким, что уже не нуждается в адекватных для своего проявления раздражителях (Lorenz 1935, 1937). Эти случаи проявления «фиксирующихся» в филогенезе реакций поведения уже граничат с настоящими инстинктами, когда определенный комплекс поведения проявляется во всем своем готовом виде, не требуя уже никакого обучения для своего проявления. По Дарвину: «Если бы трехлетний Моцарт стал играть на фортепиано не после поразительно малой практики, а совсем без практики, то справедливо было бы сказать, что он это делает инстинктивно». К группе строго фиксированных инстинктивных реакций поведения относятся многообразные, столь богато описанные в литературе инстинктивные формы поведения птиц, когда для выполнения того или другого, нередко весьма сложного действия уже не требуется индивидуального опыта, как мы это видим, например, в случае выполнения акта плавания только что вылупившимися водоплавающими птицами или акта полета молодыми ласточками. Все подобные врожденные инстинктивные реакции поведения являются уже строго наследственно фиксированными видовыми формами поведения, приспособляющими животных к тем условиям жизни, в которых в течение сотен поколений обитали их предки, борясь за свое существование как своим индивидуальным, так и видовым опытом. Таким образом, мы считаем, что индивидуально приобретенное условно-рефлекторное поведение и безусловно-рефлекторное инстинктивное поведение в своем происхождении отделены непрерывным рядом чисто количественных переходов, не позволяющих провести между ними строгой демаркационной линии. Однако это не значит, что между ними нет качественной разницы. Эта разница есть, и она чрезвычайно глубока, но ее глубина теряется, если рассматривать эти формы поведения в их историческом развитии. Можно предположить, что развитие врожденных форм поведения по пути, проложенному индивидуально приобретенными реакциями, может идти по двум направлениям эволюции нервной системы. Во-первых, различие в выработке того или другого условного рефлекса у различных особей может обусловливаться различными особенностями тех или иных подкорковых областей. В этом случае при положительном отборе особей с наибольшей наклонностью к выработке определенного условного рефлекса в филогенезе будет происходить изменение подкорки, которое в свою очередь, будет обусловливать быстроту и прочность выработки данного условного рефлекса. Это путь на закрепление и дифференцировку определенных аппаратов мозгового ствола. Сама кора головного мозга в этом случае изменяться не будет. Во-вторых, различие в выработке того или другого условного рефлекса у различных особей может обусловливаться различной функциональной активностью тех или иных систем в коре головного мозга. В этом случае при положительном отборе особей с наибольшей наклонностью к выработке определенного условного рефлекса будет происходить отбор животных с наибольшей активностью тех функциональных отделов коры головного мозга, которые связаны с выработкой этого условного рефлекса. В этом случае через отбор определенного условного рефлекса в филогенезе будет изменяться сама кора головного мозга Это путь на дифференцировку и развитие функциональных систем коры головного мозга. Мы полагаем, что филогенетическое закрепление индивидуального опыта идет по обоим указанным направлениям эволюции нервной системы. Однако первый путь наследственного закрепления индивидуального опыта через отбор соответствующих подкорковых аппаратов нам представляется более ограниченным, открывающим меньше разнообразия возможностей, чем второй. По первому пути может идти отбор индивидуально приобретенных реакций поведения, совпадающих с филогенетически уже ранее существующими у данного вида животных безусловно-рефлекторными формами поведения. Со вторым путем связано возникновение филогенетически новых реакций поведения, направление развития которых может идти в сторону приспособления коры головного мозга к легкой выработке специальных условных рефлексов, приспосабливающих животных к многообразным и новым условиям среды. Кора головного мозга, возникнув в филогенезе как орган высочайшей пластичности, великолепно приспосабливающий животное в течение онтогенеза ко всем многообразиям внешнего мира, одновременно несет в себе и черты специализации приспособления к определенным условиям жизни, выражающейся в наклонности к выработке тех или других специальных условных рефлексов. Являясь органом, ответственным в основном за возникновение временных, индивидуально приобретенных реакций поведения, кора головного мозга, быть может, способна обусловливать и некоторые черты безусловно-рефлекторного инстинктивного поведения как отражение результата наследственного фиксирования индивидуально приобретенного поведения. Морфологическое отражение этого процесса можно видеть в том, что, поднимаясь по филогенетическому ряду, кора головного мозга оказывается все более и более дифференцированной в отношении корковых проекционных зон. «Идя от птиц к более высоко организованным существам мы видим все большее и большее расхождение проекционных зон, все большее и большее проявление локализации. Добравшись до уровня наших лабораторных животных, мы видим довольно хорошо обособленные проекционные зоны, но значительно переслаивающиеся, как это себе представляют Лючиани и И. П. Павлов. Переходя к еще более высоко организованным существам, мы видим еще большее уточнение локализации» (Орбели, 1938. С. 164). Будучи, таким образом, органом, повышающим в филогенезе общую приспособляемость животных, т. е. выражением ароморфоза, по терминологии Северцова, кора головного мозга с ее функциональными центрами, возможно, является одновременно и органом, повышающем приспособляемость животных к специальным условиям жизни, т. е. способствующим идиоадаптации, по терминологии Северцова (1922). Возможные пути происхождения наследственных и инстинктивных реакций поведения, намеченные в настоящей работе, являются не единственными возможными путями возникновения данных форм поведения. Пути наследственного фиксирования индивидуально приобретенного поведения возможны только у тех животных, у которых имеются большие возможности к возникновению индивидуально приобретенного поведения. Врожденные реакции поведения, по-видимому, возникают в филогенезе не только в результате «фиксирования» индивидуальных форм поведения, но и посредством эволюции реакций поведения, связанных с низшими отделами центральной нервной системы. Это уже самостоятельный путь возникновения врожденных форм поведения у животных, стоящих на низких ступенях филогенетической лестницы и имеющих ограниченные возможности к возникновению индивидуально приобретенного поведения. Возникновение инстинктов в результате наследственного фиксирования индивидуально приобретенного поведения должно иметь у них небольшое значение. Основной путь формирования инстинктов у таких животных — это непосредственная эволюция уже существующего инстинкта. Прекрасный пример подобного рода — эволюция инстинкта строительства у паукообразных, описанная Вагнером. Как показал этот исследователь, архитектуры построек паукообразных бывают, с одной стороны, сходными у близко родственных между собой групп, с другой стороны, пауки, далеко отстоящие друг от друга в систематике, имеют постройки различного типа. Это дает возможность Вагнеру сделать вывод, что «каждая черта, каждая частность постройки иногда может быть прослежена в целом ряде генетических, связанных между собой таксономических групп». В этом случае происходит непосредственный отбор от одной инстинктивной формы поведения к другой — непосредственная эволюция самого инстинкта. Итак, мы полагаем, что филогенетическое возникновение и эволюция инстинктов может происходить: — во-первых, в результате наследственного фиксирования индивидуально приобретенных реа… Продолжение » |